Les premières étapes de l’infection par le SARS-CoV-2 sont l’interaction de Spike avec le récepteur cellulaire ACE2 suivie de la fusion entre la membrane virale et la membrane plasmique de la cellule. Le domaine S1 de Spike régit l’interaction avec ACE2 via son domaine RBD (Receptor Binding Domain) et subit plusieurs changements structuraux ainsi qu’une coupure enzymatique. Le domaine S2 peut ainsi s’ancrer dans la membrane cellulaire et induire la fusion. De plus, les cellules infectées expriment Spike à leur surface, ce qui provoque la fusion membranaire avec les cellules voisines. Ce second aspect de l’interaction Spike-ACE2 aboutit à la formation de syncitia (cellules géantes à plusieurs noyaux) associés aux nombreux dommages pulmonaires chez les cas sévères de COVID-19. La thérapie de référence est le traitement par anticorps neutralisants, qui bloquent généralement l’interaction Spike-ACE2, en ciblant le RBD.
Une équipe de l’Université de San Francisco a produit in vitro des anticorps neutralisants à partir d’échantillons génétiques de 22 patients n’ayant jamais contracté la COVID-19. Cette approche fournit des anticorps différents de ceux des patients infectés généralement étudiés. Parmi les milliards de possibilités, 6 anticorps ont été sélectionnés pour leur efficacité élevée afin de bloquer l’interaction RBD-ACE2. Malgré cette capacité commune, ces anticorps ont des pouvoirs de neutralisation variables in vitro. Si la bivalence des anticorps est supprimée (Fab monomérique), les anticorps perdent la capacité à neutraliser, ce qui montre que ceux-là ont absolument besoin de reconnaître deux sites. Par exemple, le 3D11 montre la plus forte liaison à Spike (affinité), mais la plus faible neutralisation. L’anticorps le plus puissant (5A6) peut réduire jusqu’à 10 000 fois la réplication virale et permet de maintenir l’intégrité de l’épithélium. Comme le 3D11, il se fixe sur la surface virale à une densité plus élevée. Les autres anticorps ont des propriétés intermédiaires.
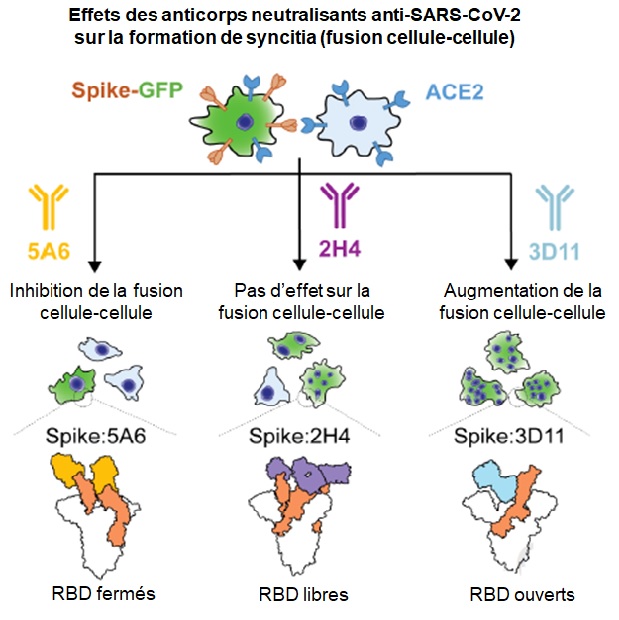
Avec un système in vitro évaluant la capacité des cellules exprimant Spike à fusionner avec d’autres cellules, les chercheurs montrent que le 5A6 bloque la formation de syncitia, alors que le 3D11 l’augmente. Les autres anticorps n’ont aucun effet sur ce mécanisme. Par cryomicroscopie électronique, ils ont déterminé que le 2H4 mime la fixation d’ACE2 et provoque chez Spike les mêmes changements structuraux qu’en présence du récepteur (ce qu’on nomme le mécanisme orthostérique), ce qui bloque la fixation d’ACE2 mais permet donc la fusion. Le 3D11 ne cible pas la zone de fixation à ACE2 (mécanisme allostérique), mais stabilise la forme finale de Spike (S2 exposé et RBD ouverts), ce qui accroît la fusion. Le 5A6 a un mode de fixation quaternaire et coopératif qui maintient les RBD en position fermée, ce qui bloque la fixation d’ACE2 et stabilise Spike sous sa forme de pré-fusion. Cette double particularité lui permet d’inhiber l’infection, mais aussi la fusion cellule-cellule.
Bien que ces 6 anticorps interagissent fortement avec le RBD de Spike (haute affinité), ils ont donc tous une bioactivité différente sur l’interaction Spike-ACE2 et la fusion. L’efficacité d’un anticorps neutralisant résulte finalement de mécanismes multifactoriels complexes et non d’une simple compétition avec ACE2 pour la fixation sur Spike.