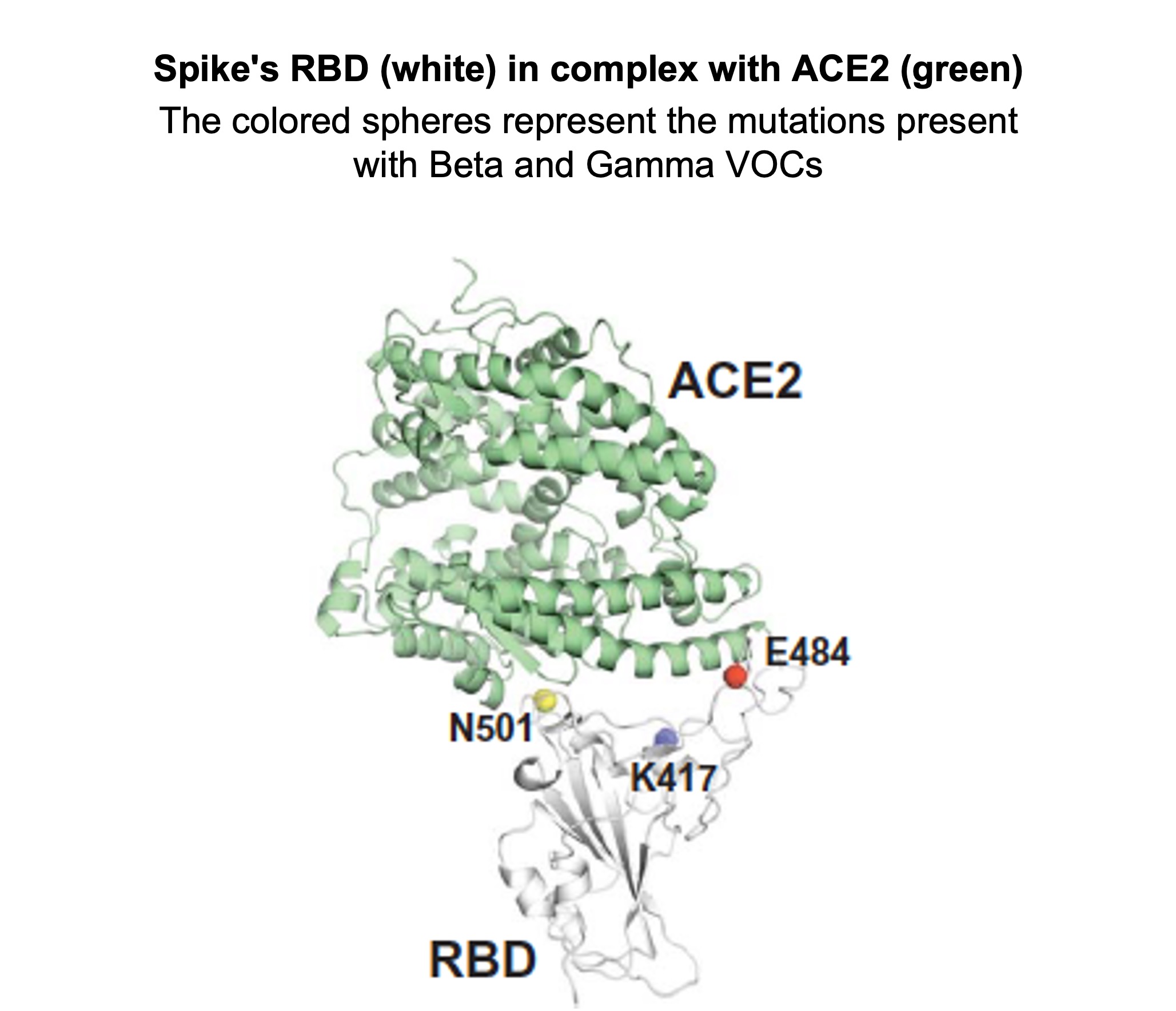
The emergence of variants of concern (VOC), different to the original Wuhan strain from both antigenic and functional points of view, call into question the efficacy of current vaccines, since they may be able to evade neutralising antibodies (nAbs). In particular, the Alpha (⍺) or UK variant, the Gamma (ɣ) or Brazilian variant, the Beta (β) or South African variant and the Epsilon (ε) or Californian variant carry mutations in the RBS (receptor binding site) of the spike. The spike is the protein that governs viral entry by interacting with the ACE2 human receptor via the RBD. This domain is the principal target of nAbs, which block the spike-ACE2 binding process. The N501Y mutation is found in the Alpha, Gamma and Beta variants, whereas the K417N/T and E484K mutations appear in the Gamma and Beta variations.
American researchers (at the Scripps Research Institute, La Jolla) have attempted to better understand the impact of these 3 mutations on the spike. They firstly established that the N501Y mutation reinforces the spike-ACE2 liaison, whereas K417N and E484K weaken it. In 585 054 SARS-CoV-2 sequences (GISAID database), 95% of K417N/T mutations were associated with N501Y (21% in all the sequences), while only 36% of E484K mutations are. The researchers therefore determined that N501Y can compensate for the loss of liaison caused by K417N, and that the triple mutant N501Y/K417N/E484K has the same effectiveness in liaison as the non-mutated protein.
The researchers then evaluated the evasion capabilities of pseudoviruses from anti-BRB nAbs derived from COVID-19 patients or humanized mice. These nAbs are representative of the germ lines most frequently enriched in anti-SARS-CoV-2 responses (IGHV3-53, IGHV3-66, IGHV1-2). The K417N and E484K mutations enable escape from several of the nAbs tested, including the most powerful IGHV3-53s, while only E484K enables evasion of IGHV1-2.
The researchers then studied the structures of 54 known anti-RBD nAbs (some of which are therapeutic). Their targets are usually classified into 6 binding sites: the RBS (RBS-A, B, C and D), the CR3022 site and the S309 site. The IGHV3-53/3-66 nAbs principally target the RBS-A and adopt a type 1 binding mode, but a minority adopt a type 2 binding mode (CDRH3 structure, with a different epitope and angle of attachment) and target the RBS-B. The majority of IGHV1-2 nAbs target the RBS-B. It appears that the N501Y and K417N affect the binding and the neutralisation of all the nAbs targeting the RBS-A, whereas the E484K affects those which target the RBS-B and -C. The L452N mutation, present in the Epsilon variant, affect the nAbs targeting the RBS-C. Amongst these antibodies, only those which interact with the RBS-D, the CR3022 and the S309 are not affected by these mutations.
The major categories of the nAbs, of which the most powerful target all the RBS-A and -B, are affected by the 3 mutations in the RBD present in the Alpha, Beta and Gamma VOC. The nAbs targeting CR3022 and S309 are in the minority and are less powerful, but are not affected by these mutations. This may explain why convalescing patients or vaccinated individuals remain partly protected against the VOC. Targeting these 2 regions may allow the development of more effective vaccines and therapies to combat the antigenic drift of SARS-CoV-2.